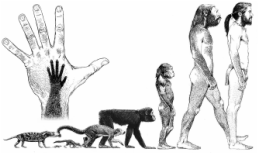
The evolution of bipedal locomotion among early hominidae is a substantially contested topic in human evolution. Numerous theories exist to explain its beginning although none are unanimously accepted. Many of the peer-reviewed journal articles pertaining to the subject employ comparative anatomy with extant primates as well as references to fossil remains of early hominidae. It is common for Australopithecus afarensis to be mentioned due to its well preserved fossil remains, and a publication date long enough ago to allow for thorough analysis and interpretation. A. afarensis displays clear cranial and postcranial evidence of bipedality, leaving the only question to be, “What were the selective pressures that led to some early hominidae developing bipedal locomotion?”
The author, Gordon W. Hewes of the University of Colorado correlates the occurrence of bipedalism with early human ancestors needing free upper extremities for food transport, “The effective use of a new food resource required its transport over considerable distances, and only bipedal locomotion, by freeing the arms and hands for carrying, could achieve maximal transportational efficiency“ (Hewes, 1961, p.689). He views the acquisition and subsequent transportation of food as more important than similar theories that involve the transport or nursing of infants, or beginnings of tool use. The infant carrying theory is refused due to the fact that current primates are fully capable of infant care via the infant clinging to the dorsal side of the mother. Tool use is rejected because of the observation that most tool use by modern humans is achieved in a sitting or squatting position. Hewes goes on to state that an increase in body weight may have been involved in the evolution of bipedalism and that the advantage of more visual range was a bi-product of it.
The authors, Elaine N. Videan and W.C. McGrew of The University of Miami in Oxford, Ohio used specific tests on chimpanzees and bonobos to observe which situation most commonly brought about an exhibition of bipedality. Assessments were for four theories on the evolution of bipedalism: carry, forage, vigilance, and display. The test for the carry theory involved the subjects being confronted with a pile of highly prized food. The subjects that used bipedal locomotion to transport the food could carry larger pieces and more total, suggesting greater efficacy of transport. This is related to the Hewes literature above because it is primarily centered on bipedalism as a result of food resources. Videan and McGrew (2002) state that, “if food availability increases the rate of bipedal locomotion other behaviors [such as tool use or infant care] will occur later at a rate greater than that of chance” in The American Journal of Physical Anthropology (p. 189).
Nina G. Jablonski and George Chaplin cite behavioral and social changes in East Africa during the middle to late Miocene as the stimulus for early hominidae to adapt to bipedalism. Dominance display rituals and appeasement behaviors are important to a species that relies on its social group for survival. Therefore, if intra- and intergroup aggression can be avoided, more individuals will be able to successfully pass on their beneficial genes. Specific bipedal postures are seen in Macaca nemestrina during threat displays and appeasement behaviors.
In addition, Jablonski and Chaplin suggest that the environmental changes that were taking place in the middle to late Miocene had a significant impact on the evolution of bipedalism. “As habitats were becoming more open and desiccated and resources more widely separated, increased intraspecific competition among pre-hominids for resources ensued“ (Jablonski & Chaplin, 1992, p. 259). Hence, the savannah grassland environment would quickly lead to the evolution of bipedal primates.
Perhaps the theory for the evolution of bipedalism that receives the least amount of support is the aquatic hypothesis. L. P. La Lumiere suggests that desiccation pushed human ancestors to the banks of the Afar triangle and stranded some on islands in the Red Sea. The extreme dryness would also have reduced the number of predators that relied on vegetation and the sea would offer protection. Bipedalism would have evolved from generations of individuals wading through shallow waters in search of food and hairlessness would have evolved for more efficient swimming. This extreme shift in environment would result in considerable evolutionary pressures that sped up the population’s adaptation for walking on two feet. Following the period of drought, the unprecedented species traveled into the Afar region of Eastern Africa by way of the lava bridge to the Ethiopian escarpment and continued to live along rivers, as it was their primary source of food and protection. L. P. La Lumiere (1981) concludes, “At the beginning of the Pleistocene, when favourable changes permitted, the upright, hominoid descendants of these apes returned to a terrestrial existence” in Philosophical Transactions of the Royal Society of London (p. 103). Where this hypothesis lacks is in its absence of tangible evidence.
An area of human anatomy where evolutionary adaptations occur in a relatively robust fashion is when offspring and reproduction are concerned. The direct relationship with the child surviving, as well as its genes, to subsequently pass those genes along once more is paramount to a species’ survival. Amitabh Avasthi (2007) stated,“Wedge-shaped vertebrae in the lower back might be the key evolutionary adaptation that helps human females maintain a stable posture over the course of pregnancy” in National Geographic News (p. 2). Fossil evidence to support this claim are apparent when the remains of Australopithecus africanus are observed. A. africanus lived approximately 2.5 million years ago, shortly after humans common ancestor with extant apes split from the genealogical tree.
Lauren Duffy, a current master’s student at Binghamton University and close friend of the author was contacted for her personal opinion on the leading selective pressure that led to human’s ancestor to become bipedal. Duffy stated that it was surely a mix of several pressures, as evolution in a complex system that concerns all variables for an individual's entire lifetime. A notable theory that has not been stated as of yet in this research was one concerning thermoregulation. A taller hominoid would have increased surface area for cooling as well as less skin in contact with the direct sunlight (vs. quadrupedalism) of the savannah where we evolved our bipedalism. Duffy concludes that while the shift from quadrupedalism to bipedalism is a gradual yet obvious shift seen in the fossil record, the factors that influenced it are much more imperceptible (L. Duffy, personal communication, July 23, 2015).
The evolutionary adaptation of bipedalism has enabled humans to become the most remarkable species on the planet. Freeing our hands for tool use or to carry food contributed to our success just as novel dominance display rituals decreased the occurrence of fatal interactions. Although the aquatic theory lacks significant evidence it cannot be ruled out as every variable must be considered. To this day no theory in unanimously accepted and the study of our beginnings persists.
References
Avasthi, A. (2007, December 12). Why Pregnant Women Don't Tip Over. Retrieved from http://news.nationalgeographic.com/news/2007/12/071212- pregnancy-tips.html
Hewes, G. W. (1961). Food Transport and the Origin of Hominid Bipedalism. American Anthropologist, 63, 687–710.
Jablonski, N. G. & Chaplin, G. (1993). Origin of habitual terrestrial bipedalism in the ancestor of the Hominidae. Journal of Human Evolution, 24, 259-280 DOI: 10.1006/jhev.1993.1021
La Lumiere, L. P. (1981). Evolution of Human Bipedalism: A Hypothesis About Where it Happened. Philosophical Transactions of the Royal Society of London, 292 (1057), 103-107.
Videan, E. N. & McGrew, W. (2002). Bipedality in chimpanzee (Pan troglodytes) and bonobo (Pan paniscus): Testing hypotheses on the evolution of bipedalism. American Journal of Physical Anthropology, 118, 184–190.
The author, Gordon W. Hewes of the University of Colorado correlates the occurrence of bipedalism with early human ancestors needing free upper extremities for food transport, “The effective use of a new food resource required its transport over considerable distances, and only bipedal locomotion, by freeing the arms and hands for carrying, could achieve maximal transportational efficiency“ (Hewes, 1961, p.689). He views the acquisition and subsequent transportation of food as more important than similar theories that involve the transport or nursing of infants, or beginnings of tool use. The infant carrying theory is refused due to the fact that current primates are fully capable of infant care via the infant clinging to the dorsal side of the mother. Tool use is rejected because of the observation that most tool use by modern humans is achieved in a sitting or squatting position. Hewes goes on to state that an increase in body weight may have been involved in the evolution of bipedalism and that the advantage of more visual range was a bi-product of it.
The authors, Elaine N. Videan and W.C. McGrew of The University of Miami in Oxford, Ohio used specific tests on chimpanzees and bonobos to observe which situation most commonly brought about an exhibition of bipedality. Assessments were for four theories on the evolution of bipedalism: carry, forage, vigilance, and display. The test for the carry theory involved the subjects being confronted with a pile of highly prized food. The subjects that used bipedal locomotion to transport the food could carry larger pieces and more total, suggesting greater efficacy of transport. This is related to the Hewes literature above because it is primarily centered on bipedalism as a result of food resources. Videan and McGrew (2002) state that, “if food availability increases the rate of bipedal locomotion other behaviors [such as tool use or infant care] will occur later at a rate greater than that of chance” in The American Journal of Physical Anthropology (p. 189).
Nina G. Jablonski and George Chaplin cite behavioral and social changes in East Africa during the middle to late Miocene as the stimulus for early hominidae to adapt to bipedalism. Dominance display rituals and appeasement behaviors are important to a species that relies on its social group for survival. Therefore, if intra- and intergroup aggression can be avoided, more individuals will be able to successfully pass on their beneficial genes. Specific bipedal postures are seen in Macaca nemestrina during threat displays and appeasement behaviors.
In addition, Jablonski and Chaplin suggest that the environmental changes that were taking place in the middle to late Miocene had a significant impact on the evolution of bipedalism. “As habitats were becoming more open and desiccated and resources more widely separated, increased intraspecific competition among pre-hominids for resources ensued“ (Jablonski & Chaplin, 1992, p. 259). Hence, the savannah grassland environment would quickly lead to the evolution of bipedal primates.
Perhaps the theory for the evolution of bipedalism that receives the least amount of support is the aquatic hypothesis. L. P. La Lumiere suggests that desiccation pushed human ancestors to the banks of the Afar triangle and stranded some on islands in the Red Sea. The extreme dryness would also have reduced the number of predators that relied on vegetation and the sea would offer protection. Bipedalism would have evolved from generations of individuals wading through shallow waters in search of food and hairlessness would have evolved for more efficient swimming. This extreme shift in environment would result in considerable evolutionary pressures that sped up the population’s adaptation for walking on two feet. Following the period of drought, the unprecedented species traveled into the Afar region of Eastern Africa by way of the lava bridge to the Ethiopian escarpment and continued to live along rivers, as it was their primary source of food and protection. L. P. La Lumiere (1981) concludes, “At the beginning of the Pleistocene, when favourable changes permitted, the upright, hominoid descendants of these apes returned to a terrestrial existence” in Philosophical Transactions of the Royal Society of London (p. 103). Where this hypothesis lacks is in its absence of tangible evidence.
An area of human anatomy where evolutionary adaptations occur in a relatively robust fashion is when offspring and reproduction are concerned. The direct relationship with the child surviving, as well as its genes, to subsequently pass those genes along once more is paramount to a species’ survival. Amitabh Avasthi (2007) stated,“Wedge-shaped vertebrae in the lower back might be the key evolutionary adaptation that helps human females maintain a stable posture over the course of pregnancy” in National Geographic News (p. 2). Fossil evidence to support this claim are apparent when the remains of Australopithecus africanus are observed. A. africanus lived approximately 2.5 million years ago, shortly after humans common ancestor with extant apes split from the genealogical tree.
Lauren Duffy, a current master’s student at Binghamton University and close friend of the author was contacted for her personal opinion on the leading selective pressure that led to human’s ancestor to become bipedal. Duffy stated that it was surely a mix of several pressures, as evolution in a complex system that concerns all variables for an individual's entire lifetime. A notable theory that has not been stated as of yet in this research was one concerning thermoregulation. A taller hominoid would have increased surface area for cooling as well as less skin in contact with the direct sunlight (vs. quadrupedalism) of the savannah where we evolved our bipedalism. Duffy concludes that while the shift from quadrupedalism to bipedalism is a gradual yet obvious shift seen in the fossil record, the factors that influenced it are much more imperceptible (L. Duffy, personal communication, July 23, 2015).
The evolutionary adaptation of bipedalism has enabled humans to become the most remarkable species on the planet. Freeing our hands for tool use or to carry food contributed to our success just as novel dominance display rituals decreased the occurrence of fatal interactions. Although the aquatic theory lacks significant evidence it cannot be ruled out as every variable must be considered. To this day no theory in unanimously accepted and the study of our beginnings persists.
References
Avasthi, A. (2007, December 12). Why Pregnant Women Don't Tip Over. Retrieved from http://news.nationalgeographic.com/news/2007/12/071212- pregnancy-tips.html
Hewes, G. W. (1961). Food Transport and the Origin of Hominid Bipedalism. American Anthropologist, 63, 687–710.
Jablonski, N. G. & Chaplin, G. (1993). Origin of habitual terrestrial bipedalism in the ancestor of the Hominidae. Journal of Human Evolution, 24, 259-280 DOI: 10.1006/jhev.1993.1021
La Lumiere, L. P. (1981). Evolution of Human Bipedalism: A Hypothesis About Where it Happened. Philosophical Transactions of the Royal Society of London, 292 (1057), 103-107.
Videan, E. N. & McGrew, W. (2002). Bipedality in chimpanzee (Pan troglodytes) and bonobo (Pan paniscus): Testing hypotheses on the evolution of bipedalism. American Journal of Physical Anthropology, 118, 184–190.